El genotipo, es el conjunto de genes que cada individuo ha heredado y que se reflejan en un determinado fenotipo o expresión de dichos genes. Este genotipo es esencialmente fijo, permanece constante a lo largo de toda la vida de un individuo y es inmodificable por efectos ambientales.
Dependiendo del medio ambiente un genotipo puede producir varios fenotipos diferentes o también varios genotipos pueden producir un solo fenotipo.
Toda la variabilidad genética de una especie, que se expresa a través de algunas características visibles en el fenotipo, pueden ser fiel reflejo del genotipo de la especie; estos caracteres son altamente heredables, presentan poca variabilidad y son poco influenciadas por el ambiente. Pero en cambio existen otros caracteres que son relevantes en la utilización de especies cultivadas, de tipo cualitativo y/o cuantitativo, que si son afectados por el ambiente y por tanto tienen poco o aceptable heredabilidad. Con la ayuda de aquellos caracteres fijos, poco influenciados por el ambiente, se puede mantener la identidad genética de una especie, lo que es sumamente importante en los procesos de fitomejoramiento.
Con la fecundación se unen los gametos, combinando dos conjuntos de genes uno de cada progenitor. Cada gen tiene una posición especifica dentro de un cromosoma que afecta a un carácter particular y está representado por dos copias, una procedente de la madre y otra del padre. Cada copia se localiza en la misma posición sobre cada uno de los cromosomas pares del cigoto.
Cuando las copias son idénticas se dice que el individuo es homocigoto para aquel gen en particular y cuando son diferentes, es decir cuando cada progenitor ha aportado una forma distinta de alelo del mismo gen, el individuo es heterocigoto para dicho gen. La forma de determinar el genotipo es a través de experimentos de reproducción o descendencia y no simplemente mediante el examen del fenotipo de un organismo.
El termino gen fue propuesto por primara vez por Mendel pero no con este nombre, sino con el termino «Factores»; estos factores eran los responsables de la transmisión de los caracteres de padres a hijos. Fue el danés Wihem L. Johansen quien acuño el termino Gen junto con los de genotipo y fenotipo e hizo clara la distinción entre genes y características de un organismo y que dichas características son el resultado de la acción de los genes.
El gen Mendeliano es una unidad de función, estructura, transmisión, mutación y evolución, que se distribuye ordenada y linealmente en los cromosomas.
A nivel genético el gen es la unidad básica de la herencia de los seres vivos y es la unidad mínima de función genética, que puede heredarse.
A nivel molecular el gen es una secuencia lineal de nucleótidos en la molécula de ADN, que contiene la información necesaria para la síntesis de una macromolécula con función celular específica. Por ejemplo: Proteínas, RNAm, RNA ribosómico, RNA de transferencia y RNA pequeños.
Los genes se encuentran en una gran molécula llamada ácido desoxirribonucleico (ADN), la cual esta asociada a una matriz de proteínas formando la cromatina o nucleoproteína y se organiza en estructuras con propiedades de tinción distintivas llamadas cromosomas que se localizan en el núcleo de la célula. La información genética fluye del ADN al ARN y de estos a las proteínas. Cada gen en un cromosoma ocupa una posición definida llamada locus.
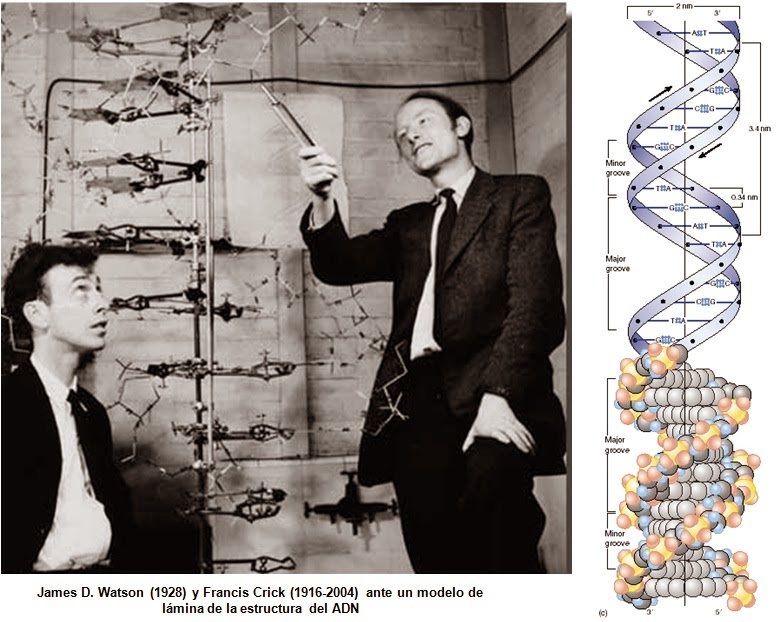
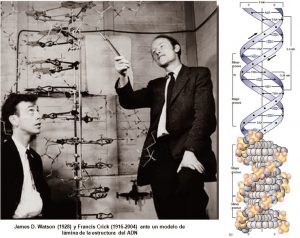
El ADN es una molécula o polímero de desoxirribonucleótidos de cadena doble, que puede ser circular o lineal, que porta la información genética con capacidad dedirigir su propia replicación y su trascripción a RNA, el cuál a su vez dirige su propia traducción a proteínas. En algunas ocasiones ocurren o pueden ocurrir cambios espontáneos o inducidos en algunas de sus partes, lo que origina una mutación la cual altera el código genético. En la figura 6 se muestra una representación esquemática de la cadena del ADN y su estructura propuesta por Watson y Crick.
Figura 6. Representación esquemática de la estructura de Watson-Crick del DNA
Como resultado de sus investigaciones con la planta de arveja (guisante), Mendel propone la teoría de la herencia particulada; con la que afirma que los caracteres están determinados por unidades genéticas discretas que se transmiten de forma intacta a través de las generaciones.
Los estudios de Mendel sentaron las bases en el sentido de que se centró en un solo carácter, en un solo fenotipo y en un modelo que además tenia varias propiedades que facilitaron explicar muchas de las observaciones que no lo eran por la teoría de la herencia mezclada, siendo muy fructífera en la comprensión del mecanismo de la herencia, tanto que se convirtieron en el prototipo del análisis genético y en los cimientos de una aproximación lógica experimental a la herencia todavía en uso.
Mendel inició sus experimentos con plantas puras de arveja, en las cuales identifico siete características distintas y contrastantes, cruzó una variedad de planta que producía semillas amarillas con otra que producía semillas verdes, estas plantas forman la Generación Parental (P).
Como resultado de este cruce salieron plantas que producían nada más que semillas amarillas, repitió los cruces con otras plantas de guisante que diferían en otros caracteres y el resultado era el mismo, salía un carácter de los dos en la generación filial. Al carácter que aparecía le llamo Dominante y al que no, Recesivo. En este caso el color amarillo es dominante frente al color verde.
Las plantas obtenidas de la Generación Parental se denominan Primera Generación Filial (F1).
Para explicarlo, Mendel concibió la idea de unas unidades hereditarias, que en la actualidad llamamos genes, los cuales expresan, a menudo, caracteres dominantes o recesivos. Al formular su primer principio (La ley de la segregación), Mendel planteó que los genes se encuentran agrupados en parejas en las células somáticas y que se segregan durante la formación de las células sexuales (gametos femeninos o masculinos). Cada miembro del par pasa a formar parte de células sexuales distintas. Cuando un gameto femenino y otro masculino se unen, se forma de nuevo una pareja de genes en la que el gen dominante (color amarillo) oculta al gen recesivo (color verde).
Para comprobar la existencia de tales unidades hereditarias Mendel dejó que se autofecundaran las plantas de la Primera Generación Filial y obtuvo la Segunda Generación Filial (F2) compuesta por plantas que producían semillas amarillas y plantas que producían semillas verdes en una proporción 3:1 (3 de semillas amarillas y 1 de semillas verdes).Repitió el experimento con otros caracteres diferenciados y obtuvo resultados similares en una proporción 3:1. Dedujo, con acierto, que los genes se agrupan en pares de los tipos AA, Aa, y aa («A» representa dominante y «a» representa recesivo).
Tras posteriores experimentos de cruzamiento, descubrió que cuando se polinizaban entre sí ejemplares AA, se producían solamente plantas de tallo alto, y que cuando los cruces se realizaban entre ejemplares aa, se obtenían sólo plantas de tallo enano. Así mismo, los cruces entre híbridos altos Aa generaban una descendencia de plantas de tallo alto y de tallo enano, en una proporción de tres a uno respectivamente. Más adelante Mendel decidió comprobar si estas leyes funcionaban en plantas diferenciadas en dos o más caracteres, eligió como Generación Parental plantas de semillas amarillas y lisas y plantas de semillas verdes y rugosas. Las cruzó y obtuvo la Primera Generacion Filial compuesta por Plantas de semillas amarillas y lisas, la primera ley se cumplía, en la F1 aparecían los caracteres dominantes (Amarillos y lisos) y no los recesivos ( Verde y rugosos ).
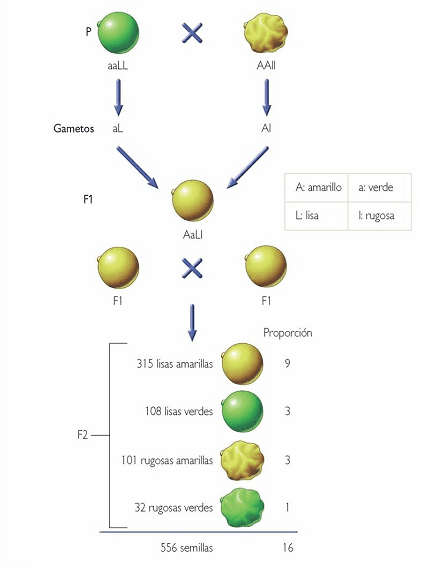
Obtuvo la Segunda Generación Filial autofecundando la Primera Generación Filial y obtuvo semillas de todos los estilos posibles, plantas que producían semillas amarillas y lisas, amarillas y rugosas, verdes y lisas y verdes y rugosas, las contó y probó con otras variedades y siempre salían en una proporción 9:3:3:1 ( 9 plantas de semillas amarillas y lisas, 3 de semillas amarillas y rugosas, 3 de semillas verdes y lisas y una planta de semillas verdes y rugosas).
Desde entonces, Mendel pudo comprender que las unidades hereditarias no se mezclan entre sí, como creían sus predecesores, sino que permanecen inalterables en el transcurso de las sucesivas generaciones. Apoyándose en esto, Mendel formuló su segundo principio (la Ley de la segregación independiente). En él se afirma que la expresión de un gen, para dar una característica física simple, no está influida, generalmente, por la expresión de otras características.
La segunda Ley o principio de la segregación resulta muy importante para el Fitomejoramiento, por que después de realizar un cruce entre dos líneas puras, en la F2 se pueden identificar caracteres que habían quedado enmascarados en la F1 y que pueden resultar provechosos para el mejoramiento y que reafirman características dominadas por genes como unidades independientes que pueden ser pasados de generación en generación. En la figura 5 se representa algunos de los resultados obtenidos por Mendel en sus experimentos con la arveja.
Figura 5 .Dihibridismo, caso de cruzamiento entre dos líneas puras o natural que difieren en 2 caracteres, segregación encontrada por Gregory Mendel. 9:3:3:1.
Las siete características con que Mendel experimento fueron:
Forma de la semilla.
Color interno de la semilla: amarillo o verde.
Color de la cubierta o tegumento de la semilla: blanco o gris.
Color de la vaina de la semilla inmadura: verde o amarilla.
Forma de la vaina de la semilla madura: Inflada o apretada.
Largo del tallo: corto (23 a 45 cm) o largo (1.50 m a 2 m)
Posición de las flores: axial (a lo largo del tallo) o Terminal (en la extremidad del tallo).
El éxito del trabajo de Mendel se debe a:
Acierto al escoger la especie: especie autogama con la que se puede obtener «individuos puros», con alta variabilidad, de diferentes colores y formas fáciles de distinguir, de ciclo corto y alta producción de semilla.
A la aplicación del método científico, que incluye observar, pensar, analizar y volver a replicar. Observación de los caracteres contrastantes, diseño cuidadoso de los experimentos y utilización del análisis estadístico, para demostrar que los resultados eran coherentes con una hipótesis explicatoria. A partir de la inferencia estadística se da una primera definición de gen y de constitución génica, para llegar a la brillante idea que los caracteres están determinados por pares de genes.
Modificación del crecimiento
y la expresión sexual en la
Cannabis por medio de
aminoetoxyvinilglicina y etefón.
·H. Y. MohanRam y Rina Sett
·Departamento de Botánica, Universidad de Delhi-110007, India
·Recibido el 13 de agosto de 1981. Aceptado el 19 de noviembre de 1981
Resumen:
La aplicación apical de aminoetoxyvinilglicina (AVG) a plantas hembra (5, 10, 25, 50 y 75 microgramos (mcg) por planta) de Cannabis sativa indujo la formación de flores machos fértiles en las recién formadas ramas primarias laterales (RPL). Se comprobó que 1 mcg por planta no es efectivo y el tratamiento con 100 mcg resultó ser inhibidor. En el caso de tratamientos desde 5 a 50 mcg,las RPL produjeron flores machos, hermafroditas y machos con fertilidad reducida, mientras que con 75 mcg formaron sólo flores machos.Cuando se aplicó AVG (25, 50 mcg) a los ápices de las plantas macho rociadas con etefón (1920 · 1-1), el efecto de feminización causado por este último se redujo notablemente.
Introducción
El etefón, un compuesto liberador de etileno (Abeles, 1973) aumenta la cantidad de flores hembra en las plantas de la familia de las cucurbitáceas (Mc Murray y Miller, 1963; Rudich et al., 1969, 1970; Augustine et al., 1973) e induce el crecimiento de flores hembra en las plantas macho de Cannabis sativa (MohanRam y Jaiswal, 1970). En dichas plantas, el ácido giberélico estimula la masculinidad (Peterson y Anhder, 1960; Wittwer y Bukovac, 1962). Se ha sugerido que la expresión sexual de la flor en la Cannabis es controlada por un balance
entre los niveles endógenos de etileno y giberélicos (MohanRam y Jaiswal, 1974). Existe evidencia de que el ion de plata es un potente inhibidor de la acción del etileno (Beyer, 1976) y que el ion de cobalto bloquea la síntesis del etileno (Beutelmann y Kende, 1977). Se ha demostrado que el nitrato de
plata y el cloruro de cobalto inducen flores macho en las plantas hembra de la Cannabis
(MohanRam y Sett, 1979; Sarath y MohanRam, 1979) y flores estaminíferas fértiles en plantas de una línea estrictamente
pistilada de Ricinuscommunis (MohanRam y Sett, 1980).
Recientemente, la aminoetoxyvinilglicina (AVG: L-a-amino-y-(2amino-ethoxy)-trans-β-ácido butanoico), un análogo estructural de la rizobitoxina ha demostrado inhibir la
producción de etileno (Bangerth, 1973; Baker et al., 1978; Amrhein y Wenker, 1979; Yu y Yang, 1979; Williams, 1980). La actual investigación se llevó a cabo para determinar el efecto de la AVG en la planta hembra de la Cannabis y también para investigar si es que el efecto del etefón puede o no ser invertido por medio de la aplicación de AVG.
Figuras 1 – 5: Cannabis sativa: 1 a. 2 a: porciones finales de una
muestra de monitores hembra y macho, respectivamente,
x0.57. – 1 b muestra una flor hembra alargada, x2.3. – 2 b es una
vista agrandada de florescencia con flores macho y brotes, x1.6.
– 3: flores modificadas de plantas hembra tratadas con AVG (50
mcg), x5. – 3 a – c son flores intersexuales, cada una con una
bráctea característica, 3 d, e son flores macho menguadas y 3 f
es una flor macho completamente alterada (ver el texto para
más detalles). – 4 a: porción final de una planta hembra tratada
con 75 mcg de AVG. Nótese que el ápice seco (flecha) y dos RLP
largas producen flores macho inducidas en las puntas (foto
tomada 48 días después del tratamiento), x0.2. – 4 b: muestra
un agrandamiento de la porción final de una de las RLP, x0.7. – 5
a: porción final de una planta macho tratado con etefón (1920 mg
● 1-1) + AVG (50 mcg). Nótese el ápice seco (flecha) y las tres RLP
que producen algunas flores intersexuales y una gran cantidad
de flores macho (mf por la sigla en inglés) (foto tomada 35 días
después del tratamiento), x0.5. – 5 b: una vista agrandada de la
punta de una RLP, x1. (br= bráctea; mf = flor macho; RLP = rama
lateral primaria; st = estigma; sta = estambre; tp = tépalo).
Material y Métodos
Almácigos de Cannabis sativa creciendo de forma natural en el Jardín Botánico de la Universidad de Delhi (en diciembre 1979,
1980), fueron trasplantados durante la etapa de 3 o 4 hojas a maceteros de arcilla llenos con tierra de jardín. Debido a que el sexo de las plantas vegetativas es no es distinguible, fue necesario esperar hasta la aparición de las primeras flores para poder separar las plantas macho y hembra. Para cada experimento se seleccionaron plantas de altura prácticamente uniforme (24 a 30 cm), que presentaban 5 o 6 nodos de floración. Se aplicó una solución acuosa de aminoetoxyvinilglicina (Hoffmann-La Roche, Nutley, Nueva Jersey, E.E.U.U.) a los ápices principales de las plantas hembra con una pipeta de 0,1 ml. Se utilizó Tween 80 -polisorbato 80- (0,01%) como tensoactivo. Se aplicó una gota de 10 mcl de la solución de prueba diariamente en un cotonete colocado en el ápice por 5 días para aplicar la cantidad total. Al final del quinto día las plantas habían recibido 1, 5, 10, 25, 50, 75 o 100 mcg de AVG.
Las muestras de monitoreo sólo se les aplicó la solución de tensoactivo (Fig. 1 a, b). A las plantas macho se les aplicó en primer lugar dos rociadas en las hojas de solución acuosa
recién preparada de 1920 mg · 1-1 de etefón (2-ácido fosfónicocloroetílico, Agromore S.A., Bangalore) con un intervalo de siete días. Posteriormente, estas plantas recibieron una aplicación apical ya sea de 25 mcg o 50 mcg de AVG un día después de la segunda rociada de etefón. Se conservaron diez plantas para cada tratamiento. Un grupo de 10 plantas sirvió como muestra de monitoreo tratadas con etefón, además de
las diez plantas macho de muestra de monitoreo que no fueron tratadas (Fig. 2 a, b) las cuales recibieron sólo la solución
tensoactiva. La viabilidad de la germinación de granos de polen en las flores macho inducidas fue puesta a prueba por medio de
germinación en un medio con 7% de sucrosa y 1% de agar o por inmersión en una solución diacetato-sucrosafluorescina (2 mg de diacetatofluorescina disueltos en una gota de 10% de solución sucrosa) mientras se observaba la fluorescencia por medio de luz ultravioleta.
Resultados
Estudios en plantas hembra
La primera respuesta visible al tratamiento con AVG (menos de 1mcg) fue la decoloración del ápice y las hojas jóvenes. Con 5 y 10 mcgel meristemo apical no se vio afectado y el crecimiento del tallo principal se reanudo después de un corto plazo. El ápice murió en el caso de dosis más elevadas (25 – 100 mcg por planta). Las plantas que recibieron 5 a 100 mcg resultaron notablemente más pequeñas que las muestras de monitoreo y la reducción de la altura dependió de la dosis aplicada. Sin
embargo, no hubo diferencia en el aumento de la cantidad de nodos posterior al tratamiento con 5 y 10 mcg. No se produjeron nuevos nodos en las plantas que recibieron 25 o más mcg, pero si se presentó cierto crecimiento en la longitud debido a
elongación intermodal.
El tratamiento con 5 a 50 mcg de AVG causó la formación de flores hembra (♀, flor con un carpelo rodeado por una bráctea grande), flores intersexuales (♀ + ♂ flor que produce tanto carpelo como estambre Fig. 3 a – c), flores macho menguado (R ♂, flor que tiene 4 o menos estambres Fig. 3 d, e), y flores macho (♂, produce cinco tépalos y cinco estambres con una copiosa cantidad de granos de polen Fig. 3 f) en las recientemente formadas ramas laterales primarias (RLP). Los nodos que se desarrollaron en el eje principal, posterior al tratamiento con 5 y 10 mcg también produjeron flores de sexo
alterado, junto con flores hembra normales.
Los datos de la Tabla 1 indican que el porcentaje de flores de sexo alterado (♀, R♂, ♂) en cada rama lateral aumenta con la
cantidad de químico aplicado. Los datos sobre la cantidad de flores se tomaron a partir de las tres primeras RLP recientemente formadas, debido a que esa cantidad fue la mínima presente en todas las plantas tratadas , con excepción de aquellas que recibieron 75mcg (en las que sólo dos RLP se desarrollaron y estas fueron más grandes que las RLP formadas en respuesta a los otros tratamientos aplicados en las muestras de monitoreo). Es nteresante acotar que en plantas tratadas con 75 mcg de AVG, no se formaron flores intersexuales o flores macho reducidas y las ramas
produjeron flores macho totalmente alteradas con unas pocas flores hembra en los nodos basales (Fig. 4 a, b).
Tabla 1: efecto de la AVG en el crecimiento del brote, largo de las ramas laterales primarias (RLP) y la expresión del sexo de las flores en plantas hembra de Cannabis1).
1) Promedio de 10 plantas, 48 días después del tratamiento; CI intervalo de certidumbre a P ≥ 0,05; D ápice seco.
2) Porcentaje de flores de sexo alterado de la cantidad total de flores presentes en cada RLP.
3) Números indican posición desde la base hacia arriba.
– Indica ausencia de flores
*) Altamente relevante sobre muestra de monitoreo a P ≥ 0,01.
NS No relevante en relación a la muestra de monitoreo.
En plantas tratadas con 100 mcg de AVG, el eje principal no produjo nuevos nodos ya que el meristemo apical murió. Las hojas jóvenes se tornaron amarillentas y quebradizas. Sin
embargo, se registró un leve aumento en la longitud de los brotes como resultado de la elongación internodal. En la mayoría de las plantas tratadas no se desarrollaron nuevas
ramas laterales. De vez en cuando, se observaron una o dos RLP, pero su crecimiento fue limitado y la producción de capullos sucedió raramente. Las flores macho inducidas produjeron granos de polen viables
y 69% de ellas germinaron in vitro a los 30-35 minutos. Del mismo modo, posterior al tratamiento con diacetato fluorescina, los granos de polen viables emitieron fluorescencia al ser observados bajo luz ultravioleta.
Etapas de la masculinización
La primera etapa hacia la “masculinidad” fue el desarrollo de una amplia gama de flores intersexuales que produjeron tanto
carpelo como estambres (Fig. 1 c). Se observó la desaparición
gradual de los órganos asociados con la flor hembra tales
como, bráctea, ovario y lóbulos del estigma. A esto le siguió la
aparición de tépalos y estambres en las flores intersexuales. La
congregación de flores en cada nodo y la presencia de
pedículos representaban características adicinales de las
plantas macho.
Se puede concluir gracias a este experimento que la dosis
mínima efectiva de AVG es de 5 mcg y la cantidad óptima es 75
mcg por planta. Al utilizar 100 mcg, la AVG produce fuerte
inhibición del crecimiento.
Estudios en plantas macho
Debido a que el etefón provoca la producción de flores hembra e intersexuales en plantas macho de Cannabis (MohanRam y Jaiswal, 1970) y la AVG induce la formación de flores macho en plantas hembra (según lo demostrado en este informe), es lógico preguntarse si es que el efecto del etefón en las plantas macho puede ser revertido por medio de la aplicación de AVG.
Se seleccionó etefón a 1920 mg ● 1-1 porque MohanRam y Jaiswal (1974) habían demostrado que una cantidad máxima de nodos (5,2) produciendo flores hembra se forma en respuesta a dicha concentración. A pesar de que se presentó una leve epinastia dos días después de la primera aplicación con espray, los efectos característicos del etefón se presentaron después de dos días de la aplicación final. Las hojas presentes a momento del tratamiento presentaron epinastia, se tornaron amarillentas y quebradizas. La extensión de la epinastia fue inversamente proporcional a la edad de la hoja. Las hojas recientemente formadas fueron más pequeñas y verduscas en comparación a las de las muestras de monitoreo no tratadas. Se presentó una importante disminución en la altura y la cantidad de nodos en todas las muestras de monitoreo tratadas en comparación a las no tratadas. Las flores macho que ya estaban presentes se resquebrajaron. Se verificó el cese del crecimiento apical en todas las plantas de muestra tratadas con etefón por un periodo de 7 a 10 días. Las RLP presentes al momento de la aplicación con espray de etefón aumentaron en tamaño, dándole a las plantas una apariencia tupida. Sin embargo, no se desarrollaron nuevas RLP inmediatamente debajo del ápice en crecimiento. En las plantas de muestra tratadas, los primeros nodos en el eje principal produjeron flores intersexuales con morfología variada, mientras que los nodos formados posteriormente produjeron sólo flores hembra. Las plantas de muestra sin tratar continuaron produciendo sólo flores macho (Tabla 2).
Tabla 2: efecto de la interacción del etefón y la AVG en el crecimiento del brote principal, tamaño de las ramas laterales primarias y la expresión sexual de la flor1)
1) Promedio de 10 plantas 35 días después del tratamiento
2) Números indican posición desde la base hacia arriba
3) Porcentaje de flores produciendo sexo alterado del total de cantidad de flores
D Indica ápice dañado
+ No hubo presencia de laterales en esta posición
CI Intervalo de certidumbre a P ≥ 0,05
*) Altamente relevante en relación a la muestra de monitoreo a P ≥ 0,01
Las plantas que recibieron tratamientos de interacción no produjeron nuevos nodos en el brote principal y el pequeño aumento en su altura fue producto de elongación internodal. Las RLP se desarrollaron después de 15 – 20 días de la inhibición del crecimiento. Por lo menos tres RLP recientemente formadas se produjeron en todas las plantas que recibieron el tratamiento de interacción. Por lo tanto los datos se tomaron sólo de las primeras tres RLP. Como respuesta al tratamiento con 25 mcg de AVG, las ramas laterales produjeron flores intersexuales, hembras y macho, aunque la cantidad de flores hembra totalmente alteradas fue menor que la cantidad de flores macho normales. Le tomo menos tiempo a las plantas volver a producir flores macho que a las muestras de monitoreo tratadas. Curiosamente, no se observaron flores hembra completamente alteradas cuando se aplicó 50 mcg de AVG a plantas tratadas con etefón. Una gran cantidad de flores macho con unas pocas flores intersexuales se formaron en las RLP (Fig. 5 a,b). La Tabla 2 muestra una drástica reducción en el porcentaje de flores alteradas. Por lo tanto el efecto feminizante de 1920 mg ● 1-1 de etefón se puede reducir notoriamente por medio de un tratamiento con 50 mcg de AVG.
Discusión
El descubrimiento de la rizobitoxina (2-amino-4-2’-amino-3’-hidroxypropoxy-trans-3-ácido butírico), una fito toxina aislada de la bacteria del nódulo de la raíz (Rhizobiumjaponicum), y capaz de inhibir la producción de etileno en tejidos de frutas y otras plantas (Owens et al., 1971), impulsaron a investigadores a buscar sus análogos estructurales. La aminoetoxivinilglicina, uno de estos análogos, había demostrado ser un efectivo inhibidor de la producción de etileno en tejidos de manzana, rebanadas de tomate verde y palta (Lieberman et al., 1974; Baker et al., 1978) y haber cortado hojas de Petunia hybrida (Gavinlertvatana et al., 1980).
La AVG ha sido utilizada para retrasar la maduración de la fruta, reducir la caída antes de la cosecha y para aumentar la firmeza de la piel, etc. (Bangerth, 1978; Williams, 1980). Sin embargo, el efecto de la AVG en la expresión sexual de la flor ha sido estudiado sólo en dos cucurbitáceas. Loy et al. (1979) informó producción mejorada de estaminífero y flores perfectas en un melón reticulado genóico. En una línea genóica de Cucumissativus indujo flores perfectas (Atsmon y Tabbak, 1979). En la presente investigación se provocaron variados grados de masculinización en las plantas hembra de Cannabis posterior a la aplicación de AVG. Curiosamente, no se desarrollaron flores intersexuales en respuesta al tratamiento con 75 mcg de AVG y las ramas laterales produjeron flores macho de manera predominantes. Se especula que la AVG gatilla expresión sexual masculina en plantas hembra al impedir la síntesis de etileno. Mientras que el ácido giberélico causa elongación del brote además de inducir la masculinización en la Cannabis sativa (MohanRam y Jaiswal, 1972), la AVG (estudio actual) retarda la elongación del brote, pero promueve la expresión sexual masculina.
Es relevante apuntar que en plantas de Cannabis macho, la AVG (50 mcg) contrarresta notablemente el efecto de dos aplicaciones de espray de 1920 mg ● 1-1 de etefón y que las RLP recientemente formadas fallan en formar flores hembra totalmente alteradas. Ness y Romani (1980) han demostrado recientemente la reversión de maduración inducida por etileno de pera por medio de AVG. Aparentemente, la AVG es capaz de superar el efecto del etefón, ya sea bloqueando su estimulación de biosíntesis de etileno o por algún otro medio hasta el momento desconocido.
Texto traducido por: Claudio Jorquera Díaz , Traductor e interprete ingles español , jorqueraucinf@gmail.com
El Cannabis afghanica ( C. sativa var. afghanica) es originario de lo que hoy es Afganistán. Es bastante baja -raramente alcanza dos metros de altura- y tiene hojas y foliolos distintivos, anchos y de color verde oscuro. Un enramado denso y entrenudos cortos, a menudo con largos rabillos para las hojas (peciolos), dominan el perfil del C. Afghanica. Los ejemplos mas comunes de C. Afghanica puro incluyen las diversas plantas de hachís existentes y las variedades afganas. El C. afghanica se cultiva exclusivamente para obtener droga, y gran parte de la resina se convierte en un hachís que es bien conocido por su alto contenido en cannabinoides. Muchos cultivadores y criadores no distinguen el C. afghanica del C. indica, y agrupan ambas variedades en la categoría del Del C. indica.
Marihuana: horticultura del cannabis la biblia del cultivador Medico de interior y exterior.
El Cannabis rudelaris ( = C. sativa var. Spontanea) fue llevada a Ámsterdam desde Europa central a principios de la década de 1980 por el seeds bank para mejorar su programa de crianza. Si no la misma rudelaris, plantas muy similares a esta variedad crecen desde el norte de Minnesota, en EE.UU., hasta Manitoba y Saskatchewan, en Canadá. El cannabis rudelaris es una planta baja y asilvestrada que contiene muy poco THC, pero empieza el ciclo de floración tras unas pocas semanas de crecimiento. El fotoperiodo no induce la floración en el C. rudelaris. Aunque a veces se confunde con plantas indicas mas potentes, el C. rudelaris puro es una autentica mala hierba que bordea los caminos, y produce dolor de cabeza en vez de embriaguez. Actualmente, unos pocos criadores han incorporado los genes de la temprana floración del C. rudelaris a otras C. sativa, C. indica y C. afganica de floración rápida.
Marihuana: horticultura del cannabis, la biblia del cultivador medico de interior y exterior. (Jorge Cervantes)
El Cannabis indica ( = C.indica var. Indica) es originario de Paquistán e india. Las variedades indica son apreciadas tanto por los cultivadores de interior como por los criadores debido a su crecimiento achaparrado y arbustivo; a su sistema de raíces condensado, los tallos robustos y las hojas anchas; y también por sus flores densas y pesadas, cargadas de THC. El follaje es un verde muy oscuro y, en algunas variedades, las hojas que rodean los cogollos se vuelven de color rojizo a púrpura. Los pistilos cortos y blanquecinos también adquieren estas tonalidades. Algunas otras C. indica de esta parte del mundo tienen hojas mas estrechas, largos pistilos blancos y follaje verde claro. Las variedades indica contienen, en general una proporción mayor de CBD por THC, lo cual provoca una efecto que es descrito a menudo como si se hubiera asentado una piedra pesada e incapacitadora en la cabeza. La potencia de la subida, varía de clara a estupefaciente. Algunas indica tienen un distinto olor, parecido al de las mofetas o la orina de gato, mientras que otras huelen dulces y exóticas. Las plantas con una abundante carga de resina tienden a ser las más resistentes a diversos hongos y plagas. Pocas indica con cogollos compactos, densos y pesados, resisten el moho gris.
Marihuana: horticultura del cannabis, la biblia del cultivador medico de interior y exterior (Jorge Cervantes)
El Cannabis sativa ( = C. sativa var. sativa) se originó predominantemente en Asia, América y África. Cada región de origen tiene características específicas, pero todas tienen los siguientes rasgos generales: gran estatura y ramas largas con espaciosas distancias internodales, un sistema de raíces ampliamente extendido, hojas grandes de foliolos estrechos, y flores un tanto escasas cuando se cultivan en interior bajo lámparas. Las variedades sativa florecen entre varias semanas y meses después que las indica. Si bien son buenas productoras al aire libre y suelen llegar a medir 4,5 m o más, las variedades sativa puras alcanzan una altura excesiva a una velocidad demasiado rápida -algunas, más de tres metros en tres meses- para resultar prácticas en el cultivo de interior. Una lámpara DAI es incapaz de iluminar plantas altas eficazmente, y el rendimiento por vatio de luz es muy bajo. Las variedades de Méjico, Colombia, Tailandia y Jamaica pueden resaltar por su potencia gracias a una proporción elevada de THC en relación al CBD, lo cual produce un efecto veloz, energético y elevador. Pero la potencia puede ser mínima también, con niveles bajos de thc. La mayoría de la marihuana que se exporta desde Colombia, Méjico, Tailandia y Jamaica recibe un trato pobre a lo largo de su vida, y aun peor durante el secado y embalaje. Estos abusos causan una degradación más rápida del THC. Como consecuencia, las semillas de este material suelen dar lugar a plantas más potentes que sus progenitoras.
Las variedades sativas de África central, incluyendo la potente congoleña, crecen de manera similar a las colombianas, con una estatura que suele superar los 4,5 metros, ramas largas y cogollos poco apretados.
Sudáfrica tiene puertos importantes y los marineros introdujeron a través de ellos el Cannabis sativa desde muchos lugares distintos. Consecuentemente, la potencia de la hierba sudafricana puede ser muy fuerte o muy débil, y las plantas pueden ser bajas, altas, de ramas largas, arbustivas, etc. La famosa Durban Poison produce potentes cogollos tempranos, de color verde claro, y es la variedad mas conocida de Sudáfrica.
El Cannabis sativa asiático incluye las variedades tailandesas, vietnamitas, laosianas, camboyanas y nepalesas, y tienen características de crecimiento diversas, así como variaciones significativas en cuanto a potencia. Al mismo tiempo que las tailandesas y otras sativas de su zona suelen ser superpotentes a base de THC, también se cuentan entre las mas difíciles de cultivar en casa y entre las mas lentas en madurar. Las variedades tailandesas producen cogollos menudos y ligeros en ramas grandes y extensas tras florecer alrededor de cuatro meses. Las sativas tailandesas, vietnamitas, camboyanas y laosianas tienen a producir adultos hermafroditas.
El Cannabis sativa nepalés puede generar hojas sobredimensionadas en planta altas, de ramas largas, que producen cogollos escasos y de floración tardía, pero otras variedades de esta región, se desarrollan como plantas bajas y compactas de floración mas rápida. Con frecuencia, la producción de THC y la potencia resultan bastantes altas, pero también suelen ser mediocres.
Todas las variedades de cáñamo se consideran Cannabis sativa. El cáñamo, al que suele llamarse cariñosamente rope (soga) en EE.UU., es C. sativa cultivado por su contenido en fibra. El cáñamo suele contener semillas y niveles apenas perceptibles de THC.
Marihuana: horticultura del cannabis, la biblia del cultivador medico de interior y exterior (Jorge Cervantes)
El cannabis ya sea soga o droga, está clasificado técnica y legalmente como cannabis sativa. Con independencia de su origen, el cannabis se considera cannabis sativa (C. sativa). Sin embargo, según Hemp diseases and pests, por el Dr. J.M. McPartland. R.C. Clarke y D. P. Watson (CAB internacional), el Cannabis Sativa puede clasificarse, además, como: Cannabis sativa, Cannabis Indica, Cannabis Rudelaris, y Cannabis Afghanica. Cada variedad tiene su propio patrón de crecimiento, así como un aspecto distinto, olor y sabor peculiares, etc.