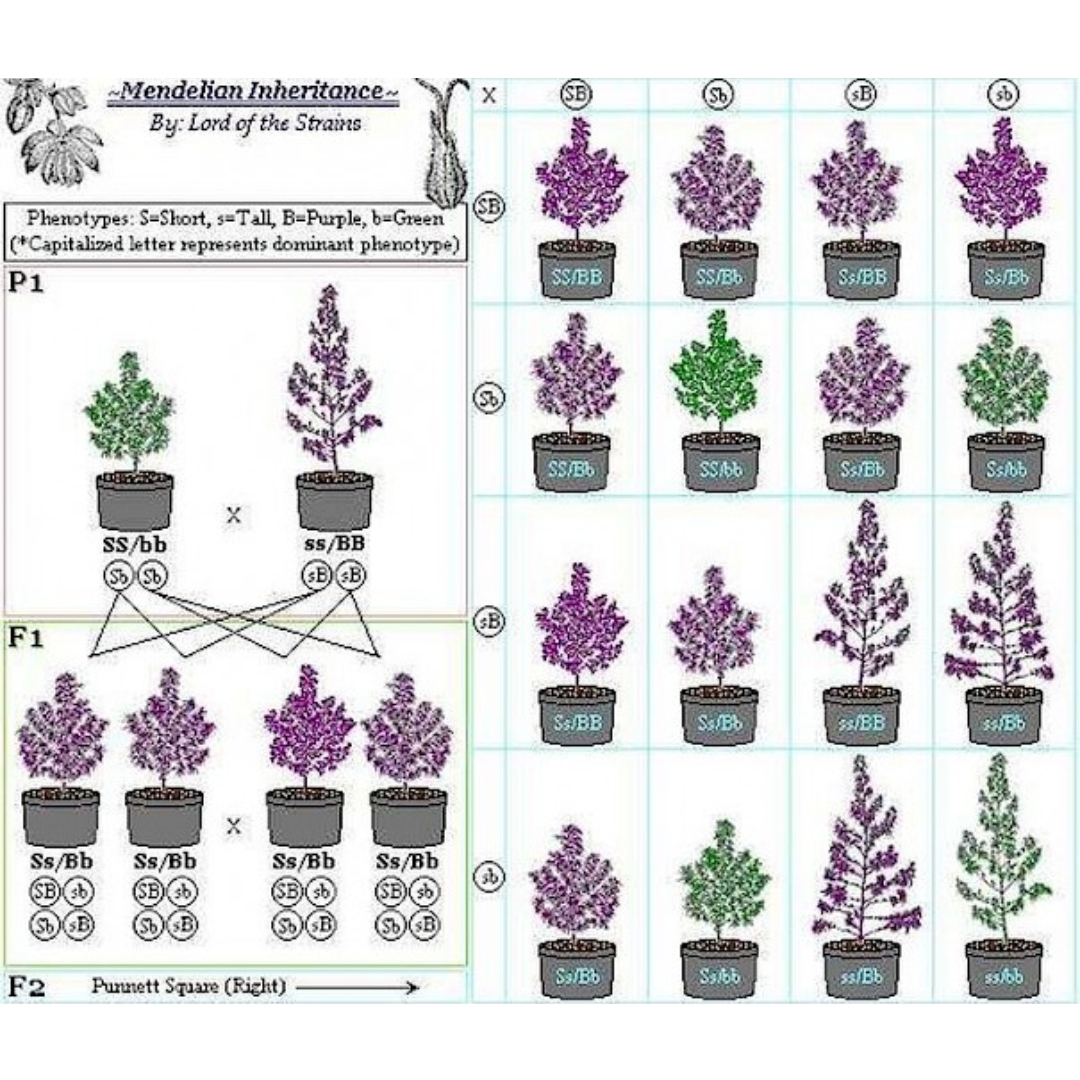
El término segregar hace referencia a apartar, separar a alguien de algo o una cosa de otra. Genéticamente segregación es la separación de cromosomas homólogos (y genes) de los diferentes progenitores en la meiosis. La segregación se hace evidente en la F2 o en generaciones posteriores de una cruza.
Cuando los miembros de la generación F1 se entrecruzaron, Mendel recobro muchos descendientes con semillas amarillas, y algunos de semillas verdes. Luego del análisis estadístico de la generación F2, Mendel determinó que la relación entre plantas con semillas amarillas/verdes era 3:1. Las plantas con semillas verdes no aparecían en la primera generación F1, y se encontraban en la segunda F2 y sucesivas generaciones.
Mendel concluyó que el carácter estudiado estaba gobernado por factores discretos (separables) y que el rasgo del carácter que aparece en la F1 es el dominante. Los factores se heredaban a pares, teniendo cada generación un par de los mismos. Actualmente nos referimos a esos factores como alelos. El hecho de que los caracteres se hereden de a pares permiten explicar el fenómeno observado del «salto» de una generación.
El cruzamiento en las plantas se realiza mediante la reproducción sexual de dos individuos diferentes, dando como resultado una descendencia que hereda parte del material genético de cada progenitor. Los organismos parientes deben ser genéticamente compatibles y pueden ser de variedades diferentes o de especies muy cercanas. El cruzamiento originalmente se da bajo polinización cruzada natural entre plantas cuya constitución genética es diferente.
El cruzamiento en las plantas, como en los animales, se da como una forma de evitar la endogamia y aumentar al máximo la diversidad genética y las oportunidades asociadas para la supervivencia.
El cruzamiento entre variedades o entre especies relativamente distantes es un método muy utilizado en el mejoramiento de la productividad de las plantas. Para ello fue necesario conocer la forma como se heredaban las características de los progenitores e identificar los caracteres deseables.
El cruzamiento tiene entonces como objetivos: combinar alelos, introducir alelos, seleccionar alelos en la obtención de materiales o líneas mejoradas.
Mendel fue el primero que estudió la herencia, iniciando con la forma de la semilla de la arveja. Él cruzó una raza pura de plantas de semillas lisas con una raza pura de otra que siempre producía semillas rugosas (60 fertilizaciones en 15 plantas). Todas las semillas resultantes resultaron lisas. A este fenómeno se le llamó Cruzamiento de un solo carácter o monohibridación.
Al año siguiente, Mendel plantó esas semillas y permitió que las mismas se auto fecundasen. Recogió 7324 semillas en total: 5474 lisas y 1850 rugosas. Para sistematizar el registro de datos, las generaciones fueron nombradas y numeradas. La generación parental se denomina como P. Los descendientes de la generación P son la generación F1 (la primera filial). La autofecundación de la generación de F1 produce la generación F2 (la segunda filial).
P1. Lisa X Rugosa
F1 Todas lisas
F2. 5474 lisas y 1850 rugosas
Lo mismo sucedió con cada par de caracteres elegidos: cuando cepas puras de plantas con semillas amarillas se cruzaron con razas puras de plantas con semillas verdes, todos los descendientes fueron plantas con semillas amarillas. Los padres del entrecruzamiento son la generación P1, y los descendientes representan la generación F1.
Manual para la producción de Semillas feminizadas con tiosulfato de plata. STS
Se sugiere trabajar la feminización con plantas obtenidas desde semilla regular y hacer la selección de una planta mayormente pistilifera (hembra) que no muestre rasgos de intersexualidad (hermafroditismo)
Se deben considerar dos cámaras de cultivo, una para el proceso vegetativo y una para la floración, en la cámara de crecimiento a: 18/6 se mantendrán las plantas madres.
El ideal es que sean dos variedades de distinta condición genética por ej: Sativa/Indica (Brasil x Sur de la India) es el cruce original de White Widow, también puede ser entre híbridos por citar algunos: Jack47 (Jack Herer x AK47); Jackwidow (Jack Herer x White widow)
Por la condición del cannabis de pertenecer a la familia Alogama-Dioica le favorece la polinización cruzada. Una ves realizado el trabajo de selección de los parentales y para no perder el avance el ideal es trabajar la reversión del sexo con esquejes.
Ya que las plantas no reaccionan de manera automática a una concentración de sts estándar, se debe hacer una reversión de prueba para cada esqueje y para esto es muy importante numerar los esquejes con la disolución aplicada
Se debe realizar todo el proceso en un cuarto oscuro ya que el nitrato de plata es fotosensible, se debe iluminar únicamente con una ampolleta bombilla o foco LED, que de luz de color rojo: NO se debe utilizar NINGUN INSTRUMENTO METALICO.
Se deben utilizar medidas de prevención al manipular el producto. El nitrato de plata produce quemaduras en la piel en forma de manchas oscuras, no duele pero las manchas permanecen durante varias semanas, aunque nunca es bueno que nuestra piel tenga contacto con productos químicos, ni siquiera con los productos de limpieza del hogar.
Las plantas tratadas y los restos de productos no aplicados, deben considerarse como productos contaminantes. No existe información sobre el consumo de las plantas tratadas; deben ser desechadas.
Solución 1 Nitrato de Plata 10 ml. Solución 2 tiosulfato de sodio 40 ml. Estas dos soluciones se deben guardar por separado en lugar oscuro y a baja temperatura hasta el momento de su uso. También se necesitara 200ml de agua destilada. Para completar 250ml de sts base.
Preparación: Agregar la solución 1 nitrato de plata (10 ml) a los 200 ml de agua destilada de manera muy lenta sin precipitar, después se agrega la solución 2 tiosulfato de sodio (40 ml) también muy lentamente sin precipitar mejor si cada mezcla se hace gota a gota, obtendremos en total 250ml de STS BASE.
Una vez mezcladas las soluciones 1 y 2, solo tiene estabilidad a corto plazo pero de todas maneras servirá para una segunda aplicación, se debe mantener refrigerado pero después de la segunda aplicación no es conveniente guardarlo.
Los 250ml de STS BASE se deben diluir para ser utilizado se sugiere hacerlo en 500ml de agua destilada (1:2) en la reversion de prueba puedes aplicar distintas concentraciones
Antes de aplicar el sts por el anverso y reverso de las hojas se debe tapar el sustrato con un plástico para que no llegue a las raíces, una ves aplicado el sts se pasa a la cámara de floración con el foco y ventiladores apagados hasta el día siguiente, hasta cuando se prenda el foco a un fotoperiodo de 12/12.
Hay quienes aplican nuevamente despues 7,10,15 ó 20 días una aplicación también es suficiente eso dependerá de cada variedad y se resolverá en la reversión de prueba. Recuerda que después de aplicar el sts la planta debe permanecer a oscuras y con los ventiladores apagados hasta que el producto se ha fijado y secado (una noche es suficiente).
entre 20 y 30 días después de la aplicación comenzaran a aparecer las primeras flores macho (poliniferas) en cuanto notes que están apareciendo introduces los clones receptores que serán fecundados con el polen revertido.
este punto es muy importante ya que si introduces los esquejes de los parentales al mismo tiempo a la cámara de floración puede que en los esquejes receptores su proceso de floración este muy avanzado y las semillas no alcancen a madurar completamente este proceso también dependerá de la rapidez de la floraciónde cada variedad que es un punto importante a considerar.
Nota: Se deben utilizar solo los elementos que van incluidos en el kit sts, ya que si se usa otra botella u otros utensilios que no estén lo suficientemente limpios, puede estropear la mezcla y por ende su efectividad.
Se consideran plantas autógamas a aquellas que utilizan la autofecundación como mecanismo reproductivo.
Se considera como alógamas a aquellas plantas que no se auto fecundan, sino que, por el contrario, poseen mecanismos de fecundación cruzada.
La alogamia es un sistema que garantiza la variabilidad genética y por tanto las nuevas combinaciones alélicas dentro de una especie. En la evolución de plantas fueron apareciendo mecanismos de reproducción que favorecían la alogamia y que excluían total o parcialmente la autogamia. Las ventajas de la alogamia radican en la producción de nuevas combinaciones genéticas en la población, que aseguran la variabilidad de la especie y, en consecuencia, la posibilidad de sobrevivir a los cambios de medio ambiente. Por eso las Angiospermas desarrollaron numerosas adaptaciones florales para favorecer la alogamia, como por ejemplo la separación espacial y temporal de los sexos y otras variaciones como la presentación secundaria de polen.
Según el tipo de flor que posean, las plantas alógamas se clasifican en tres grupos:
Plantas con flores hermafroditas: Son flores completas que poseen los dos sexos. Ejemplo de ellas son: Cebolla, el centeno y el maracuyá.
Plantas monoicas: Tienen flores unisexuales masculinas y femeninas en la misma planta, como el melón, el mijo, el pepino y el maíz.
Plantas dioicas: Plantas con flores masculinas y plantas con flores femeninas, como araucaria y kiwi.
Para una especie alógama o de fecundación cruzada, el proceso de autofecundación o cruzamientos con individuos estrechamente emparentados, lleva a grandes problemas con consecuencias drásticas para algunos alelos pues durante el proceso de autofecundación se manifiestan alelos letales que no se manifestaban en el estado heterocigótico.
En las poblaciones de plantas alógamas, es frecuente la presencia de factores desfavorables que se mantienen en heterocigosis, tales como genes de esterilidad floral en centeno, letales por deficiencias clorofílicas en maíces y otros. En la mayoría de los casos estos genes se mantienen porque la selección natural favorece a los heterocigotos.
Los genes considerados letales hacen que el individuo no llegue a desarrollarse, muriendo en estado de embrión. Sin embargo, a veces la letalidad no es completa y se considera más bien como subletalidad, caracterizada por una disminución de la sobre vivencia de los individuos homocigóticos para un determinado gen o por la manifestación a una determinada edad de los efectos del gen.
Las plantas alógamas han desarrollado mecanismos que les permiten mantener o incentivar la alogamia, tales como la dicogamia y la presencia de barreras mecánicas.
Plantas dioicas
Las especies dioicas son plantas bisexuales. Cada especie presenta individuos con flores masculinas e individuos con flores femeninas, lo que determina la alogamia obligada. Esto significa que las dos estructuras reproductoras se hallan en plantas diferentes (plantas estaminadas y plantas pistiladas).
Muchas plantas dioicas presentan flores relativamente pequeñas, blancas, amarillentas o verdosas, de morfología no especializada, que atraen una variedad de insectos pequeños.
La dioecia o diferenciación sexual es claramente un mecanismo de fecundación cruzada que impide la autofecundación, pero no impide el apareamiento entre hermanos o formas más estrechas de consanguinidad.
A menudo la dioecia está asociada con plantas de gran tamaño y polinización abiótica. Es rara en plantas con flores grandes, especializadas, con morfología compleja. Entre las plantas cultivadas las especies dioicas más importantes son: la palmera africana, cáñamo, lúpulo, espinaca, papayo, la marihuana, álamo y el espárrago.
El fenotipo es la manifestación visible del genotipo en un determinado ambiente, por lo tanto, está determinado fundamentalmente por el genotipo o por la identidad de los alelos, los cuales, individualmente, cargan una o más posiciones en los cromosomas. Algunos fenotipos están determinados por los múltiples genes y además influenciados por factores del medio. De esta manera, la identidad de uno o de unos pocos alelos conocidos, no siempre permite una predicción del fenotipo. En este sentido, la interacción entre el genotipo y el fenotipo ha sido descrita usando siguiente ecuación: Fenotipo = Genotipo + Ambiente
La constitución genotípica de cada planta depende de cierta manera de la forma en que ésta se reproduce, por lo tanto, el sistema reproductivo de cada planta estará relacionado con el método específico que debe utilizarse para poder mejorarla.
Las plantas que se reproducen sexualmente mediante polinización pueden hacerlo de varías formas, ya sea fertilizándose a sí mismas o fertilizándose unas a otras entre individuos de la misma especie. Así, será importante determinar qué tipo de fertilización se da en cada especie a fin de poder manipular prácticas como las de polinización artificial antes de comenzar programas de mejora genética.
El genotipo, es el conjunto de genes que cada individuo ha heredado y que se reflejan en un determinado fenotipo o expresión de dichos genes. Este genotipo es esencialmente fijo, permanece constante a lo largo de toda la vida de un individuo y es inmodificable por efectos ambientales.
Dependiendo del medio ambiente un genotipo puede producir varios fenotipos diferentes o también varios genotipos pueden producir un solo fenotipo.
Toda la variabilidad genética de una especie, que se expresa a través de algunas características visibles en el fenotipo, pueden ser fiel reflejo del genotipo de la especie; estos caracteres son altamente heredables, presentan poca variabilidad y son poco influenciadas por el ambiente. Pero en cambio existen otros caracteres que son relevantes en la utilización de especies cultivadas, de tipo cualitativo y/o cuantitativo, que si son afectados por el ambiente y por tanto tienen poco o aceptable heredabilidad. Con la ayuda de aquellos caracteres fijos, poco influenciados por el ambiente, se puede mantener la identidad genética de una especie, lo que es sumamente importante en los procesos de fitomejoramiento.
Con la fecundación se unen los gametos, combinando dos conjuntos de genes uno de cada progenitor. Cada gen tiene una posición especifica dentro de un cromosoma que afecta a un carácter particular y está representado por dos copias, una procedente de la madre y otra del padre. Cada copia se localiza en la misma posición sobre cada uno de los cromosomas pares del cigoto.
Cuando las copias son idénticas se dice que el individuo es homocigoto para aquel gen en particular y cuando son diferentes, es decir cuando cada progenitor ha aportado una forma distinta de alelo del mismo gen, el individuo es heterocigoto para dicho gen. La forma de determinar el genotipo es a través de experimentos de reproducción o descendencia y no simplemente mediante el examen del fenotipo de un organismo.
El termino gen fue propuesto por primara vez por Mendel pero no con este nombre, sino con el termino «Factores»; estos factores eran los responsables de la transmisión de los caracteres de padres a hijos. Fue el danés Wihem L. Johansen quien acuño el termino Gen junto con los de genotipo y fenotipo e hizo clara la distinción entre genes y características de un organismo y que dichas características son el resultado de la acción de los genes.
El gen Mendeliano es una unidad de función, estructura, transmisión, mutación y evolución, que se distribuye ordenada y linealmente en los cromosomas.
A nivel genético el gen es la unidad básica de la herencia de los seres vivos y es la unidad mínima de función genética, que puede heredarse.
A nivel molecular el gen es una secuencia lineal de nucleótidos en la molécula de ADN, que contiene la información necesaria para la síntesis de una macromolécula con función celular específica. Por ejemplo: Proteínas, RNAm, RNA ribosómico, RNA de transferencia y RNA pequeños.
Los genes se encuentran en una gran molécula llamada ácido desoxirribonucleico (ADN), la cual esta asociada a una matriz de proteínas formando la cromatina o nucleoproteína y se organiza en estructuras con propiedades de tinción distintivas llamadas cromosomas que se localizan en el núcleo de la célula. La información genética fluye del ADN al ARN y de estos a las proteínas. Cada gen en un cromosoma ocupa una posición definida llamada locus.
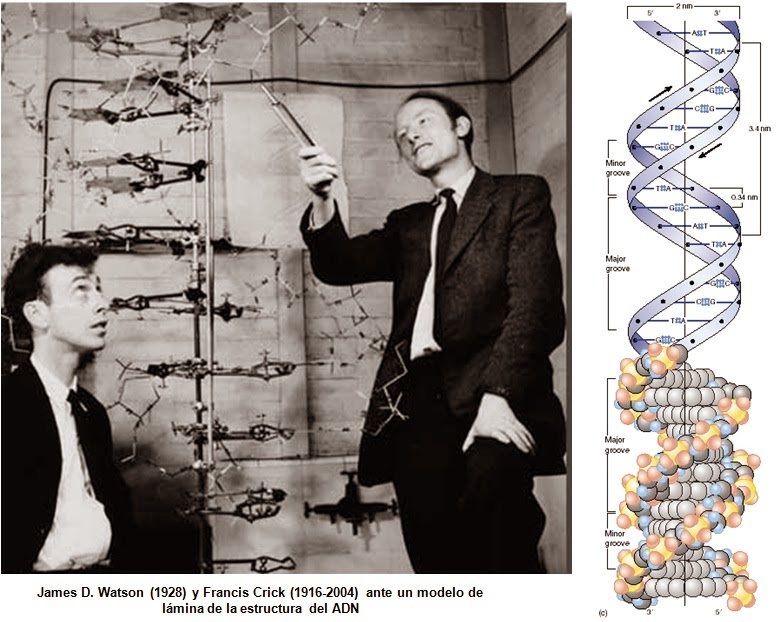
El ADN es una molécula o polímero de desoxirribonucleótidos de cadena doble, que puede ser circular o lineal, que porta la información genética con capacidad dedirigir su propia replicación y su trascripción a RNA, el cuál a su vez dirige su propia traducción a proteínas. En algunas ocasiones ocurren o pueden ocurrir cambios espontáneos o inducidos en algunas de sus partes, lo que origina una mutación la cual altera el código genético. En la figura 6 se muestra una representación esquemática de la cadena del ADN y su estructura propuesta por Watson y Crick.
Figura 6. Representación esquemática de la estructura de Watson-Crick del DNA
Modificación del crecimiento
y la expresión sexual en la
Cannabis por medio de
aminoetoxyvinilglicina y etefón.
·H. Y. MohanRam y Rina Sett
·Departamento de Botánica, Universidad de Delhi-110007, India
·Recibido el 13 de agosto de 1981. Aceptado el 19 de noviembre de 1981
Resumen:
La aplicación apical de aminoetoxyvinilglicina (AVG) a plantas hembra (5, 10, 25, 50 y 75 microgramos (mcg) por planta) de Cannabis sativa indujo la formación de flores machos fértiles en las recién formadas ramas primarias laterales (RPL). Se comprobó que 1 mcg por planta no es efectivo y el tratamiento con 100 mcg resultó ser inhibidor. En el caso de tratamientos desde 5 a 50 mcg,las RPL produjeron flores machos, hermafroditas y machos con fertilidad reducida, mientras que con 75 mcg formaron sólo flores machos.Cuando se aplicó AVG (25, 50 mcg) a los ápices de las plantas macho rociadas con etefón (1920 · 1-1), el efecto de feminización causado por este último se redujo notablemente.
Introducción
El etefón, un compuesto liberador de etileno (Abeles, 1973) aumenta la cantidad de flores hembra en las plantas de la familia de las cucurbitáceas (Mc Murray y Miller, 1963; Rudich et al., 1969, 1970; Augustine et al., 1973) e induce el crecimiento de flores hembra en las plantas macho de Cannabis sativa (MohanRam y Jaiswal, 1970). En dichas plantas, el ácido giberélico estimula la masculinidad (Peterson y Anhder, 1960; Wittwer y Bukovac, 1962). Se ha sugerido que la expresión sexual de la flor en la Cannabis es controlada por un balance
entre los niveles endógenos de etileno y giberélicos (MohanRam y Jaiswal, 1974). Existe evidencia de que el ion de plata es un potente inhibidor de la acción del etileno (Beyer, 1976) y que el ion de cobalto bloquea la síntesis del etileno (Beutelmann y Kende, 1977). Se ha demostrado que el nitrato de
plata y el cloruro de cobalto inducen flores macho en las plantas hembra de la Cannabis
(MohanRam y Sett, 1979; Sarath y MohanRam, 1979) y flores estaminíferas fértiles en plantas de una línea estrictamente
pistilada de Ricinuscommunis (MohanRam y Sett, 1980).
Recientemente, la aminoetoxyvinilglicina (AVG: L-a-amino-y-(2amino-ethoxy)-trans-β-ácido butanoico), un análogo estructural de la rizobitoxina ha demostrado inhibir la
producción de etileno (Bangerth, 1973; Baker et al., 1978; Amrhein y Wenker, 1979; Yu y Yang, 1979; Williams, 1980). La actual investigación se llevó a cabo para determinar el efecto de la AVG en la planta hembra de la Cannabis y también para investigar si es que el efecto del etefón puede o no ser invertido por medio de la aplicación de AVG.
Figuras 1 – 5: Cannabis sativa: 1 a. 2 a: porciones finales de una
muestra de monitores hembra y macho, respectivamente,
x0.57. – 1 b muestra una flor hembra alargada, x2.3. – 2 b es una
vista agrandada de florescencia con flores macho y brotes, x1.6.
– 3: flores modificadas de plantas hembra tratadas con AVG (50
mcg), x5. – 3 a – c son flores intersexuales, cada una con una
bráctea característica, 3 d, e son flores macho menguadas y 3 f
es una flor macho completamente alterada (ver el texto para
más detalles). – 4 a: porción final de una planta hembra tratada
con 75 mcg de AVG. Nótese que el ápice seco (flecha) y dos RLP
largas producen flores macho inducidas en las puntas (foto
tomada 48 días después del tratamiento), x0.2. – 4 b: muestra
un agrandamiento de la porción final de una de las RLP, x0.7. – 5
a: porción final de una planta macho tratado con etefón (1920 mg
● 1-1) + AVG (50 mcg). Nótese el ápice seco (flecha) y las tres RLP
que producen algunas flores intersexuales y una gran cantidad
de flores macho (mf por la sigla en inglés) (foto tomada 35 días
después del tratamiento), x0.5. – 5 b: una vista agrandada de la
punta de una RLP, x1. (br= bráctea; mf = flor macho; RLP = rama
lateral primaria; st = estigma; sta = estambre; tp = tépalo).
Material y Métodos
Almácigos de Cannabis sativa creciendo de forma natural en el Jardín Botánico de la Universidad de Delhi (en diciembre 1979,
1980), fueron trasplantados durante la etapa de 3 o 4 hojas a maceteros de arcilla llenos con tierra de jardín. Debido a que el sexo de las plantas vegetativas es no es distinguible, fue necesario esperar hasta la aparición de las primeras flores para poder separar las plantas macho y hembra. Para cada experimento se seleccionaron plantas de altura prácticamente uniforme (24 a 30 cm), que presentaban 5 o 6 nodos de floración. Se aplicó una solución acuosa de aminoetoxyvinilglicina (Hoffmann-La Roche, Nutley, Nueva Jersey, E.E.U.U.) a los ápices principales de las plantas hembra con una pipeta de 0,1 ml. Se utilizó Tween 80 -polisorbato 80- (0,01%) como tensoactivo. Se aplicó una gota de 10 mcl de la solución de prueba diariamente en un cotonete colocado en el ápice por 5 días para aplicar la cantidad total. Al final del quinto día las plantas habían recibido 1, 5, 10, 25, 50, 75 o 100 mcg de AVG.
Las muestras de monitoreo sólo se les aplicó la solución de tensoactivo (Fig. 1 a, b). A las plantas macho se les aplicó en primer lugar dos rociadas en las hojas de solución acuosa
recién preparada de 1920 mg · 1-1 de etefón (2-ácido fosfónicocloroetílico, Agromore S.A., Bangalore) con un intervalo de siete días. Posteriormente, estas plantas recibieron una aplicación apical ya sea de 25 mcg o 50 mcg de AVG un día después de la segunda rociada de etefón. Se conservaron diez plantas para cada tratamiento. Un grupo de 10 plantas sirvió como muestra de monitoreo tratadas con etefón, además de
las diez plantas macho de muestra de monitoreo que no fueron tratadas (Fig. 2 a, b) las cuales recibieron sólo la solución
tensoactiva. La viabilidad de la germinación de granos de polen en las flores macho inducidas fue puesta a prueba por medio de
germinación en un medio con 7% de sucrosa y 1% de agar o por inmersión en una solución diacetato-sucrosafluorescina (2 mg de diacetatofluorescina disueltos en una gota de 10% de solución sucrosa) mientras se observaba la fluorescencia por medio de luz ultravioleta.
Resultados
Estudios en plantas hembra
La primera respuesta visible al tratamiento con AVG (menos de 1mcg) fue la decoloración del ápice y las hojas jóvenes. Con 5 y 10 mcgel meristemo apical no se vio afectado y el crecimiento del tallo principal se reanudo después de un corto plazo. El ápice murió en el caso de dosis más elevadas (25 – 100 mcg por planta). Las plantas que recibieron 5 a 100 mcg resultaron notablemente más pequeñas que las muestras de monitoreo y la reducción de la altura dependió de la dosis aplicada. Sin
embargo, no hubo diferencia en el aumento de la cantidad de nodos posterior al tratamiento con 5 y 10 mcg. No se produjeron nuevos nodos en las plantas que recibieron 25 o más mcg, pero si se presentó cierto crecimiento en la longitud debido a
elongación intermodal.
El tratamiento con 5 a 50 mcg de AVG causó la formación de flores hembra (♀, flor con un carpelo rodeado por una bráctea grande), flores intersexuales (♀ + ♂ flor que produce tanto carpelo como estambre Fig. 3 a – c), flores macho menguado (R ♂, flor que tiene 4 o menos estambres Fig. 3 d, e), y flores macho (♂, produce cinco tépalos y cinco estambres con una copiosa cantidad de granos de polen Fig. 3 f) en las recientemente formadas ramas laterales primarias (RLP). Los nodos que se desarrollaron en el eje principal, posterior al tratamiento con 5 y 10 mcg también produjeron flores de sexo
alterado, junto con flores hembra normales.
Los datos de la Tabla 1 indican que el porcentaje de flores de sexo alterado (♀, R♂, ♂) en cada rama lateral aumenta con la
cantidad de químico aplicado. Los datos sobre la cantidad de flores se tomaron a partir de las tres primeras RLP recientemente formadas, debido a que esa cantidad fue la mínima presente en todas las plantas tratadas , con excepción de aquellas que recibieron 75mcg (en las que sólo dos RLP se desarrollaron y estas fueron más grandes que las RLP formadas en respuesta a los otros tratamientos aplicados en las muestras de monitoreo). Es nteresante acotar que en plantas tratadas con 75 mcg de AVG, no se formaron flores intersexuales o flores macho reducidas y las ramas
produjeron flores macho totalmente alteradas con unas pocas flores hembra en los nodos basales (Fig. 4 a, b).
Tabla 1: efecto de la AVG en el crecimiento del brote, largo de las ramas laterales primarias (RLP) y la expresión del sexo de las flores en plantas hembra de Cannabis1).
1) Promedio de 10 plantas, 48 días después del tratamiento; CI intervalo de certidumbre a P ≥ 0,05; D ápice seco.
2) Porcentaje de flores de sexo alterado de la cantidad total de flores presentes en cada RLP.
3) Números indican posición desde la base hacia arriba.
– Indica ausencia de flores
*) Altamente relevante sobre muestra de monitoreo a P ≥ 0,01.
NS No relevante en relación a la muestra de monitoreo.
En plantas tratadas con 100 mcg de AVG, el eje principal no produjo nuevos nodos ya que el meristemo apical murió. Las hojas jóvenes se tornaron amarillentas y quebradizas. Sin
embargo, se registró un leve aumento en la longitud de los brotes como resultado de la elongación internodal. En la mayoría de las plantas tratadas no se desarrollaron nuevas
ramas laterales. De vez en cuando, se observaron una o dos RLP, pero su crecimiento fue limitado y la producción de capullos sucedió raramente. Las flores macho inducidas produjeron granos de polen viables
y 69% de ellas germinaron in vitro a los 30-35 minutos. Del mismo modo, posterior al tratamiento con diacetato fluorescina, los granos de polen viables emitieron fluorescencia al ser observados bajo luz ultravioleta.
Etapas de la masculinización
La primera etapa hacia la “masculinidad” fue el desarrollo de una amplia gama de flores intersexuales que produjeron tanto
carpelo como estambres (Fig. 1 c). Se observó la desaparición
gradual de los órganos asociados con la flor hembra tales
como, bráctea, ovario y lóbulos del estigma. A esto le siguió la
aparición de tépalos y estambres en las flores intersexuales. La
congregación de flores en cada nodo y la presencia de
pedículos representaban características adicinales de las
plantas macho.
Se puede concluir gracias a este experimento que la dosis
mínima efectiva de AVG es de 5 mcg y la cantidad óptima es 75
mcg por planta. Al utilizar 100 mcg, la AVG produce fuerte
inhibición del crecimiento.
Estudios en plantas macho
Debido a que el etefón provoca la producción de flores hembra e intersexuales en plantas macho de Cannabis (MohanRam y Jaiswal, 1970) y la AVG induce la formación de flores macho en plantas hembra (según lo demostrado en este informe), es lógico preguntarse si es que el efecto del etefón en las plantas macho puede ser revertido por medio de la aplicación de AVG.
Se seleccionó etefón a 1920 mg ● 1-1 porque MohanRam y Jaiswal (1974) habían demostrado que una cantidad máxima de nodos (5,2) produciendo flores hembra se forma en respuesta a dicha concentración. A pesar de que se presentó una leve epinastia dos días después de la primera aplicación con espray, los efectos característicos del etefón se presentaron después de dos días de la aplicación final. Las hojas presentes a momento del tratamiento presentaron epinastia, se tornaron amarillentas y quebradizas. La extensión de la epinastia fue inversamente proporcional a la edad de la hoja. Las hojas recientemente formadas fueron más pequeñas y verduscas en comparación a las de las muestras de monitoreo no tratadas. Se presentó una importante disminución en la altura y la cantidad de nodos en todas las muestras de monitoreo tratadas en comparación a las no tratadas. Las flores macho que ya estaban presentes se resquebrajaron. Se verificó el cese del crecimiento apical en todas las plantas de muestra tratadas con etefón por un periodo de 7 a 10 días. Las RLP presentes al momento de la aplicación con espray de etefón aumentaron en tamaño, dándole a las plantas una apariencia tupida. Sin embargo, no se desarrollaron nuevas RLP inmediatamente debajo del ápice en crecimiento. En las plantas de muestra tratadas, los primeros nodos en el eje principal produjeron flores intersexuales con morfología variada, mientras que los nodos formados posteriormente produjeron sólo flores hembra. Las plantas de muestra sin tratar continuaron produciendo sólo flores macho (Tabla 2).
Tabla 2: efecto de la interacción del etefón y la AVG en el crecimiento del brote principal, tamaño de las ramas laterales primarias y la expresión sexual de la flor1)
1) Promedio de 10 plantas 35 días después del tratamiento
2) Números indican posición desde la base hacia arriba
3) Porcentaje de flores produciendo sexo alterado del total de cantidad de flores
D Indica ápice dañado
+ No hubo presencia de laterales en esta posición
CI Intervalo de certidumbre a P ≥ 0,05
*) Altamente relevante en relación a la muestra de monitoreo a P ≥ 0,01
Las plantas que recibieron tratamientos de interacción no produjeron nuevos nodos en el brote principal y el pequeño aumento en su altura fue producto de elongación internodal. Las RLP se desarrollaron después de 15 – 20 días de la inhibición del crecimiento. Por lo menos tres RLP recientemente formadas se produjeron en todas las plantas que recibieron el tratamiento de interacción. Por lo tanto los datos se tomaron sólo de las primeras tres RLP. Como respuesta al tratamiento con 25 mcg de AVG, las ramas laterales produjeron flores intersexuales, hembras y macho, aunque la cantidad de flores hembra totalmente alteradas fue menor que la cantidad de flores macho normales. Le tomo menos tiempo a las plantas volver a producir flores macho que a las muestras de monitoreo tratadas. Curiosamente, no se observaron flores hembra completamente alteradas cuando se aplicó 50 mcg de AVG a plantas tratadas con etefón. Una gran cantidad de flores macho con unas pocas flores intersexuales se formaron en las RLP (Fig. 5 a,b). La Tabla 2 muestra una drástica reducción en el porcentaje de flores alteradas. Por lo tanto el efecto feminizante de 1920 mg ● 1-1 de etefón se puede reducir notoriamente por medio de un tratamiento con 50 mcg de AVG.
Discusión
El descubrimiento de la rizobitoxina (2-amino-4-2’-amino-3’-hidroxypropoxy-trans-3-ácido butírico), una fito toxina aislada de la bacteria del nódulo de la raíz (Rhizobiumjaponicum), y capaz de inhibir la producción de etileno en tejidos de frutas y otras plantas (Owens et al., 1971), impulsaron a investigadores a buscar sus análogos estructurales. La aminoetoxivinilglicina, uno de estos análogos, había demostrado ser un efectivo inhibidor de la producción de etileno en tejidos de manzana, rebanadas de tomate verde y palta (Lieberman et al., 1974; Baker et al., 1978) y haber cortado hojas de Petunia hybrida (Gavinlertvatana et al., 1980).
La AVG ha sido utilizada para retrasar la maduración de la fruta, reducir la caída antes de la cosecha y para aumentar la firmeza de la piel, etc. (Bangerth, 1978; Williams, 1980). Sin embargo, el efecto de la AVG en la expresión sexual de la flor ha sido estudiado sólo en dos cucurbitáceas. Loy et al. (1979) informó producción mejorada de estaminífero y flores perfectas en un melón reticulado genóico. En una línea genóica de Cucumissativus indujo flores perfectas (Atsmon y Tabbak, 1979). En la presente investigación se provocaron variados grados de masculinización en las plantas hembra de Cannabis posterior a la aplicación de AVG. Curiosamente, no se desarrollaron flores intersexuales en respuesta al tratamiento con 75 mcg de AVG y las ramas laterales produjeron flores macho de manera predominantes. Se especula que la AVG gatilla expresión sexual masculina en plantas hembra al impedir la síntesis de etileno. Mientras que el ácido giberélico causa elongación del brote además de inducir la masculinización en la Cannabis sativa (MohanRam y Jaiswal, 1972), la AVG (estudio actual) retarda la elongación del brote, pero promueve la expresión sexual masculina.
Es relevante apuntar que en plantas de Cannabis macho, la AVG (50 mcg) contrarresta notablemente el efecto de dos aplicaciones de espray de 1920 mg ● 1-1 de etefón y que las RLP recientemente formadas fallan en formar flores hembra totalmente alteradas. Ness y Romani (1980) han demostrado recientemente la reversión de maduración inducida por etileno de pera por medio de AVG. Aparentemente, la AVG es capaz de superar el efecto del etefón, ya sea bloqueando su estimulación de biosíntesis de etileno o por algún otro medio hasta el momento desconocido.
Texto traducido por: Claudio Jorquera Díaz , Traductor e interprete ingles español , jorqueraucinf@gmail.com